Metabolism
1.1 Introduction
In vertebrates, the primary organ to metabolize chemicals is the liver, the major portal entry of xenobiotics. In other taxa, such as molluscs and arthropods, other organs, such as digestive gland (molluscs) and hepatopancreas (arthropods) display a key role in chemical´s metabolism. By altering the dose of a chemical or by adding another molecule to it, xenobiotics-metabolizing enzymes can alter the toxicity of the compound, its tissue-binding properties, as well as its distribution and persistence within the body (Figure 1).
In a toxicological perspective, the basic function of metabolism is to increase the rate of elimination of a xenobiotic. Metabolism involves a process of biotransformation in which the chemical structure of the xenobiotic is changed. This change increases the water solubility of the chemical, which results in a reduction of the chemical ability to be stored in the adipose tissue and an increment of its elimination rate.
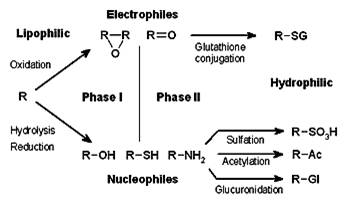
Figure 1- Chemical biotransformation.
Source: http://upload.wikimedia.org/wikipedia/commons/thumb/9/97/Xenobiotic_metabolism.png/350px-Xenobiotic_metabolism.png
Although the main benefit of
biotransformation is to protect the
organism from high chemical levels within the different
tissues, the lack of specificity and predictability of the process can
lead to a bioactivation. However, enzymatic bioactivation of many
environmental
chemicals is a necessary step in the process of detoxification and
elimination of the xenobiotic. Depending on the chemical nature of the
toxic substance, detoxification can take several pathways and may
compete with bioactivation/toxication of a chemical. Interindividual
variance between organisms may account for the variation in the
response to
chemical exposure and the extent or type of observed toxic effects.
Another important determinant of metabolizing capacities is the prior
exposure to chemicals that can potentially induce or inhibit enzymes
involved in the process.
To illustrate this unit, see the following video:
https://www.youtube.com/watch?v=ztsBn8gsfHw
1.2 Phase I and Phase II reactions
Metabolism requires the contribution of different xenobiotic-metabolizing enzymes, which occur at the main locations of possible chemical exposure, namely the lungs, gastrointestinal track, skin and eyes, being present as well in the kidneys, many endocrine glands, the brain and the red blood cells. The liver contains the highest concentration of these enzymes, which is consistent with its primary function as receptor of the majority of the xenobiotics absorbed by the organism. The bacteria present in the gastrointestinal track also present important metabolizing enzymes that enable the organism to metabolize a variety of chemicals. Moreover, just as some xenobiotics are biotransformed by the endobiotic-metabolizing enzymes in intermediate metabolism processes, certain endobiotics are biotransformed by the xenobiotic-metabolizing enzymes.
The liver is specialized in the synthesis of many important biologically functional proteins and is also capable of mediating chemical transformations of several xenobiotics. Generally, these xenobiotic compounds are lipophilic, whereby they are likely to bind to lipid membranes and accumulate in adipose tissue as well as to be transported by lipoproteins in the blood and reach different compartments/organs. Within the liver, hepatocytes or parenchymal cells are the major site of drug biotransformation. The highly vascular nature of the tissue and the intimate contact between blood and hepatocytes, which contain the xenobiotic-metabolizing enzymes, allows for the rapid diffusion of chemicals in and metabolites out. The xenobiotic-metabolizing enzymes carry out chemical modifications on foreign chemicals, which make easier the process of elimination from the body.
While most enzymes are very specific to their substrate, xenobiotic-metabolizing enzymes are able to metabolize different substrates due to their unusual plasticity, allowing stretching and distortion. Indeed, the active site of some of these enzymes can accommodate multiple substrates. There may be more than one metabolic reaction at a single site and more than one enzyme/isozyme capable of catalyzing a single reaction at a single site (Table 1).
Table 1 - General pathways of xenobiotic biotransformation, enzymes involved and their major subcellular location.
| Reaction | Enzyme or specific reaction | Localization |
|---|---|---|
| Hydrolysis | Carboxylesterase | Microsomes, cytosol, lysosomes, blood |
| Alkaline phosphatase | Plasma membrane | |
| Peptidase | Blood, lysosomes | |
| Epoxide hydrolase | Microsomes, cytosol | |
| Reduction | Azo- nitro-reduction | Microflora |
| Carbonyl (aldo-keto) reduction | Cytosol, microsomes, blood | |
| Disulfide reduction | Cytosol | |
| Sulfoxide reduction | Cytosol | |
| Quinone reduction | Cytosol, microsomes | |
| Dihydropyrimidine dehydrogenase | Cytosol | |
| Reductive dehalogenation | Microsomes | |
| Dehydroxylation (cytochrome b5) | Microsomes | |
| Dehydroxylation (aldehyde oxidase) | Cytosol | |
| Oxidation | Alcohol dehydrogenase | Cytosol |
| Aldehyde dehydrogenase | Mitochondria, cytosol | |
| Aldehyde oxidase | Cytosol | |
| Xanthine oxidase | Mitochondria | |
| Monoamine oxidase | Mitochondria | |
| Diamine oxidase | Cytosol | |
| Peroxidase | Microsomes, lysosomes, saliva | |
| Flavin-monooxygenases | Microsomes | |
| Cytochrome P450 | Microsomes | |
| Conjugation | UDP-Glucuronosyltransferase | Microsomes |
| Sulfotransferase | Cytosol | |
| Glutathione transferase | Cytosol, microsomes, mitochondria | |
| Amino acid transferase | Mitochondria, microsomes | |
| N-Acetyltransferase | Mitochondria, cytosol | |
| Methyltransferase | Cytosol, microsomes, blood |
Source: Klaassen CD. (2008)
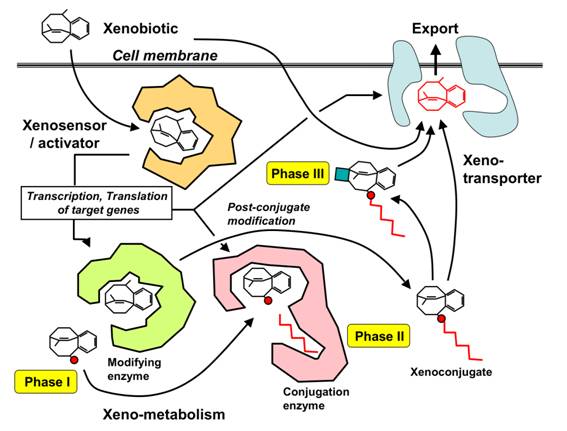
Figure 2 - Scheme of the chemical biotransformation process (Phases I, II and III).
Source: http://biopolyverse.files.wordpress.com/2012/03/xenorecoglevels.png
Although not a portal of entry, the kidney is an organ where
xenobiotics are likely to be concentrated during the excretion process
and this may explain the high level of xenobiotic-metabolizing enzymes
in this tissue.
In many cases, the toxicity of a xenobiotic is due to the parent compound, in which case xenobiotic biotransformation serves as a detoxication mechanism, by modifying the biological properties of the compound. Although this biotransformation can make the xenobiotic less toxic, in some cases the secondary metabolite is more toxic than the parental compound (bioactivation). Moreover, xenobiotic-metabolizing-enzymes can convert certain xenobiotics into reactive metabolites (e.g., electrophiles and radicals) and this activation process plays an important role in chemical cellular toxicity and chemicals mutagenicity or carcinogenicity, including oxidative stress. The balance between activation and detoxification by xenobiotic-metabolizing enzymes is often a key determinant of chemical toxicity, and often the basis for organ or species differences in toxicity.
1.2.1 Phase I
Oxidations
Microsomal oxidations are predominantly catalyzed by a group of enzymes called mixed-function oxidases or monooxygenases but also a flavoprotein. The terminal oxidase is generally a hemoprotein called cytochrome P450 (commonly abbreviated CYP or simply P450 enzymes), arguably the most highly researched family of proteins. Typically, oxidation proceeds by adding oxygen to a foreign compound (e.g., hydroxyl group insertion in benzene to form phenol) or, alternatively, by removing hydrogen atoms from a molecule (e.g., ethanol from alcoholic beverages is converted to acetaldehyde by alcohol dehydrogenase) (Figure 3).
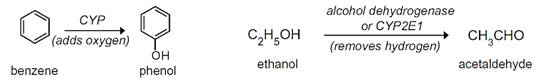
Figure 3 - Benzene and Ethanol oxidation reactions.
Source: Burcham, PC. (2014)
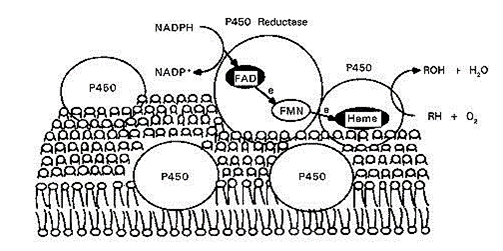
Figure 4 - Oxidation reaction and electron transporter chain.
Source: https://encrypted-tbn2.gstatic.com/images?q=tbn:ANd9GcSvrr6cWMhxZunDNIxbTBXn1tRR9y-82gJ5gJI54gZbScgJ65k3
Flavoprotein-catalyzed oxidations differ from cytochrome
P450-catalyzed oxidations in mechanism and in substrate selectivity. In
this oxidation reaction, the enzyme forms an activated oxygen complex
which is discharged in addition of a metabolized chemical (Figure 5).
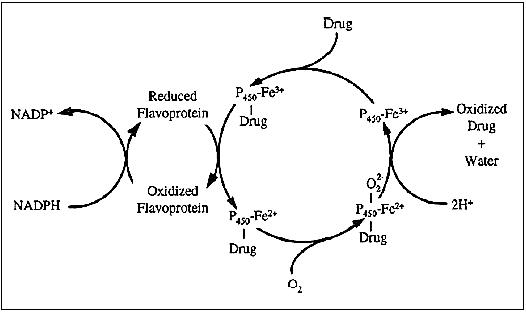
Figure 5 - Flavoprotein-catalyzed oxidation.
Source: http://www.biology-online.org/user_files/Image/Biochemistry/Enzymology/Cytochrome%20P450%20f01.jpg
Nonmicrosomal oxidations in other subcellular organelles can be flavoproteins or pyridine nucleotide linked dehydrogenases.
| Alcohol Dehydrogenase | |
| Aldehyde Dehydrogenase | |
|
Amine oxidases - monoamine oxidases - diamine oxidases |
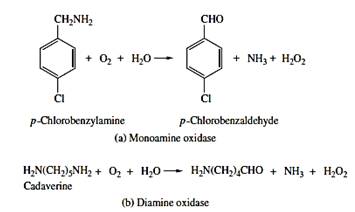 |
Source: Hodgson E. (2010)
Hydrolyses
Hydrolyses reaction are catalyzed by esterases and amidases (Figure 6). While both can be microsomal, esterases are predominantly located in cytosol.
Epoxide hydrolyses activity is predominantly microsomal, but this enzyme is also present in the cytosol. Most hydrolyses occur to a significant extent in tissues other than liver.
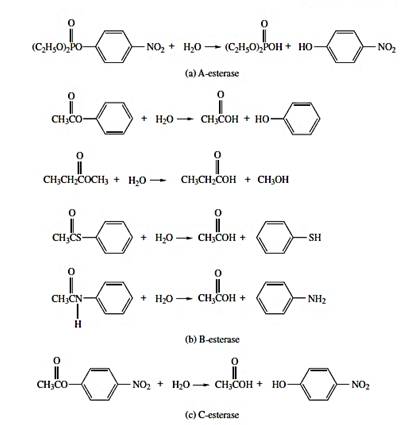
Figure 6 - Examples of esterase/amidase reactions involving xenobiotics.
Source: Hodgson, E. (2010).
Reduction
Reductive metabolism occurs in the endoplasmatic reticulum of hepatocytes through the mediation of both hemoprotein (cytochrome P450) and flavoproteins. The most frequent reactions are the reductions of azo- and nitro-groups, but other reductions occur namely, disulfides, sulfoxides, epoxides and N-oxides. Under aerobic conditions, the products of reductive metabolism can be reoxidized.
This biotransformation involves the addition of extra hydrogen atoms to a molecule (Figure 7) or the removal of oxygen atoms.
Figure 7 - Reduction reactions by addition of extra hydrogen atoms.
Source: Burcham PC. (2014).
1.2.2 Phase II
Products of Phase I metabolism and other xenobiotics containing functional groups such as hydroxyl, amino, carboxyl, epoxide or halogen can undergo conjugation reactions with endogenous metabolites in phase II reactions. Generally, conjugation products are more polar, less toxic and more readily excreted than are their parent compounds.
Glucuronidation
The glucuronidation reaction is one of the major pathways for elimination of many lipophilic xenobiotics and endobiotics from the body. This biotransformation is catalyzed by UDP glucuronosyltransferases (UGT), located in the endoplasmatic reticulum, which transfer a glucuronic acid from a uridine 5 2-diphosphoglucuronic acid (UDPGA) cofactor to a functional group or phase I metabolite, forming a glucuronide conjugate (Figure 8).
| A | 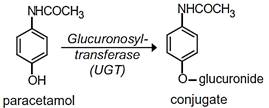 |
| B | 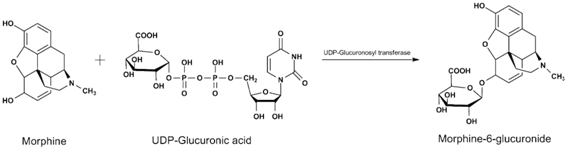 |
| C | 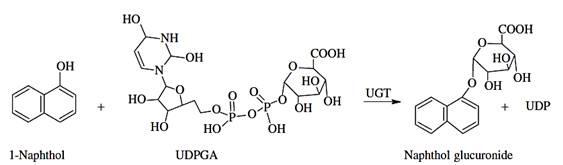 |
Figure 8 - Examples of glucoronidation reactions.
Source: (A) Burcham PC. (2014); (B) http://www.covachem.com/blog/wp-content/uploads/2014/09/Glucuronidation-Reaction.png; (C) Hodgson E. (2010).
In addition to competing with CYP for the active site of some
foreign chemicals, UGT proteins metabolize also many products of
CYP-catalyzed reactions.
Glucoside conjugation
Although rare in vertebrates, glucosides formed from xenobiotics are
common in insects and plants. Formed from UDP-glucose, they appear to
fall into the same classes as the glucuronides.
Sulfonation
Besides the limited sulfate availability within the cell, sulfate conjugation is an important alternative to glucuronidation for phenolic compounds and occasionally arylamines. Sulfonation reactions are catalyzed by sulfotransferases (SULT) and involve the transfer of a sulfonate group (-SO3) from a cofactor (PAPS - 3'-phosphoadenosine-5'-phosphosulfate) onto a nucleophilic xenobiotic. Since their substrate preferences frequently overlap, UGT and SULT proteins often compete for the same xenobiotic substrate (Figure 9).
| A | 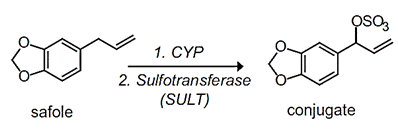 |
| B | 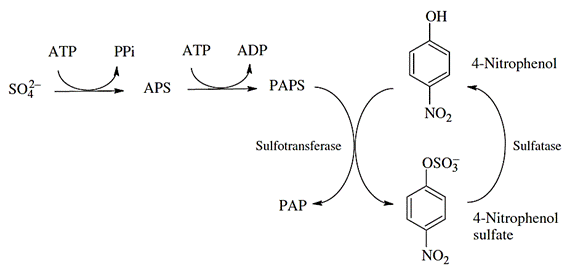 |
Source: (A) Burcham, PC. (2014); (B) Hodgson E. (2010).
The SULT enzymes are expressed strongly in the GIT, liver, kidneys,
platelets, brain and unlike many other biotransformation genes, SULT
genes are expressed in fetal tissues.
Glutathione conjugation
Glutathione is a small peptide comprising only three amino acids (glutamate, cysteine and glycine) and in addition to its importance in xenobiotic metabolism, it is also involved in important reactions for sequestering toxic metabolites generated by biotransformation of some chemicals.
Although small peptides are rapidly degraded by peptidases within most cells, the unusual γ-carboxyl linkage between glutamate and cysteine contribute to glutathione resistance to proteolysis.
Under normal physiological conditions, glutathione reductase (GR) maintains glutathione predominantly within the reduced form (GSH). In addition to direct oxidation of GSH, glutathione disulfide (GSSG) is also formed during enzymatic detoxication of xenobiotics and inorganic or organic peroxides by glutathione peroxidise (GP) (Figure 10). Glutathione conjugation is accelerated by glutathione-S-transferases (GSTs), a ubiquitous enzyme family found in cytosolic, microsomal and mitochondrial cells fractions and that confers cell protection against endogenous electrophiles and reactive intermediates formed during xenobiotic metabolism.
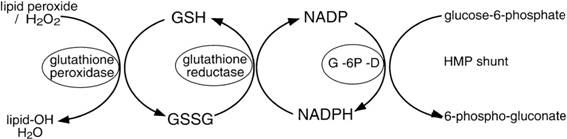
Figure 10 - Scheme of glutathione conjugation reactions.
Source: http://d2swrb3wp8hjrz.cloudfront.net/content/ajplung/277/6/L1067/F2.large.jpg
Acylation
Acylation reactions are of two general types, acetylation and amino acid conjugation. The conjugations involving acetylation of xenobiotics containing sulfonamide or amine groups and peptide conjugation of xenobiotics containing carboxylic groups do not contribute significantly to an enhanced excretion through an increase in water solubility.
Both reactions involve the activated conjugated agent coenzyme A (CoA) as a cofactor and require the formation of a thioester with the carboxylic group, either of acetate, or of the xenobiotic. The thioester then reacts with an amine, either on the xenobiotic (acetylation) or amino acid (amino acid conjugation).
Acetylation is catalyzed by the cytosolic enzymes N-acetyltransferases (Figure 11a). Amino acid conjugation reaction, exogenous carboxylic acids are activated to form S-CoA derivatives in a reaction involving ATP and CoA (Figure 11 b). These CoA derivatives then acylate the amino group of a variety of amino acids. Glycine and glutamate appear to be the most common acceptor of amino acids in mammals. In reptiles and birds the amino acid ornithine assumes that function whereas in fish the most common acceptor of amino acid is taurine.
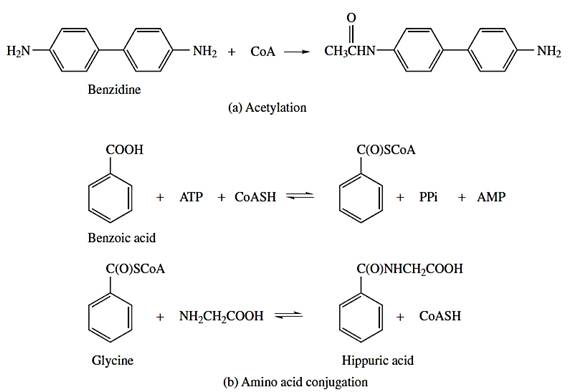
Figure 11 - Examples of acylation reactions. (a) Acetylation; (b) Amino acid conjugation.
Source: Hodgson E. (2010)
Methylation
Similarly to acylation reactions, methylation of xenobiotics containing amine and catechol groups do not play a significant role in biotransformation/detoxification process. A large number of both endogenous and exogenous compounds can be methylated by several N-, O-, and S- methyltransferases. Methylations reactions require the formation of S-adenosylmethionine (SAM) from ATP and the amino acid, methionine (Figure 12).
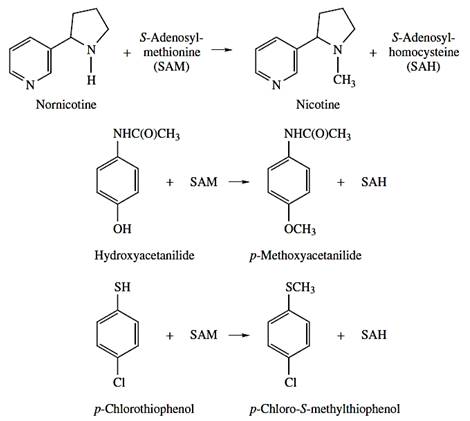
Figure 12 - Examples of methyltransferase reactions.
Source: Hodgson E. (2010)
1.3 ABC transporters
Adenosine-triphosphate-binding cassette (ABC) transporters constitute a large superfamily of both importers and exporters membrane proteins. These ATP-dependent protein complexes mediate transport of several compounds across membranes by hydrolysis of the phosphate bond between the γ- and the β-phosphate of ATP. Then, the free energy released after ATP conversion to ADP and orthophosphate can be used to accumulate the transported substrates, controlling their movement between cellular compartments or intracellular/extracellular environments by modification of protein conformation. The side of the membrane where ATP is bound and hydrolyzed is termed cis-side, whereas the opposite is trans-side.
These proteins were first described as biochemical factors conferring multidrug resistance (MDR) in cancer. ABC transporters also show high expression levels in tissues involved in excretion/detoxification (e.g., liver and kidneys), or acting as barriers (gut epithelium, capillary endothelia forming the blood-brains barrier, epithelial cells...) mediating the cellular efflux of a diverse array of chemicals and some metals and limiting chemical uptake. In analogy to the MDR phenomenon in cancer cells, Kurelec and collegues adopted the term "multixenobiotic resistance (MXR) proteins" for ABC drug efflux transport in aquatic animals, emphasizing the biochemical protective functions of the cellular pumps against environmental pollution. However, there are currently some concerns about possible negative effects of several environmental chemicals in the activity of these ABC xenobiotic transporters. An inhibition of transport function of these pumps could lead to an increased sensitivity to xenobiotics, termed chemosensitisation.
The molecular structure of all ABC transporters includes two transmembrane (TM) domains (TMDs) or subunits and two nucleotide-binding domains (NBDs) (Figure 13). However, the term "ABC transporter" is only used when the NBDs form a complex with TMDs. In contrast to the TMDs, the NBDs are highly conserved and are not always associated with TMDs, as they may also be involved in various functions that do not occur at the membrane.
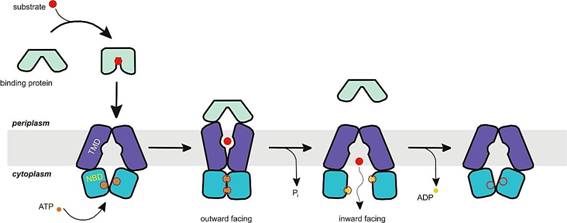
Source: http://upload.wikimedia.org/wikipedia/commons/d/d1/Abc_importer.jpg
ABC transporters with import function have a high-affinity binding
protein (BP) that specifically associates with the substrate in the
periplasm for delivery to the appropriate ABC transporter. Exporters do
not have the BP. Instead, these complexes have an intracellular domain
(ICD) that joins the membrane-spanning helices and the ABC complex
domain. It is believed that ICD is responsible for communication
between the TMD and NBD.